(от греч. méiosis - уменьшение)
редукционное
деление, деления созревания, способ деления клеток, в результате которого происходит уменьшение (редукция) числа хромосом в два раза и одна диплоидная
клетка (содержащая два набора хромосом) после двух быстро следующих друг за другом делений даёт начало 4 гаплоидным (содержащим по одному набору хромосом). Восстановление диплоидного числа хромосом происходит в результате оплодотворения. М. - обязательное звено полового процесса и условие формирования половых клеток (гамет (См.
Гаметы)). Биологическое значение М. заключается в поддержании постоянства
Кариотипа в ряду поколений организмов данного вида и обеспечении возможности рекомбинации (См.
Рекомбинация) хромосом и генов при половом процессе. М. - один из ключевых механизмов наследственности (См.
Наследственность) и наследственной изменчивости (См.
Изменчивость). Поведение хромосом при М. обеспечивает выполнение основных законов наследственности (см.
Менделя законы).
В зависимости от места М. в жизненном цикле (См.
Жизненный цикл) организмов различают 3 типа М. Гаметный, или терминальный, М. (у всех многоклеточных животных и ряда низших растений), происходит в половых органах и приводит к образованию гамет. Зиготный, или начальный, М. (у многих грибов и водорослей), происходит в зиготе (См.
Зигота) сразу после оплодотворения и приводит к образованию гаплоидного мицелия (См.
Мицелий) или
Таллома, а затем спор и гамет. Споровый, или промежуточный, М. (у высших растений), имеет место накануне цветения и приводит к образованию гаплоидного
Гаметофита, в котором позднее образуются гаметы. У простейших (Protozoa) встречаются все 3 типа М. Перед М. происходит удвоение количества ДНК в клетке. В ходе двух делений М. (
рис.) ДНК делится поровну между 4 клетками. В результате первого (редукционного) деления М. пары гомологичных хромосом разъединяются и члены пар расходятся в 2 клетки (редукция числа хромосом). Каждая хромосома сохраняет две продольные половины - хроматиды (См.
Хроматида). В результате второго (эквационного) деления хроматиды расходятся в разные клетки и каждая из 4 сестринских клеток получает по одной хроматиде. Т. о., первое
деление М. принципиально отличается от
Митоза, а второе - это
митоз в клетках с гаплоидным числом хромосом. Во время М., перед редукцией числа хромосом, происходит обмен участками гомологичных хромосом -
Кроссинговер, приводящий к перераспределению аллельных генов (см.
Аллели). М. длится много дольше митоза: например, у пшеницы он продолжается 24 часа, у лилии - 9-12 суток, у мыши - 11-14 суток, у человека - 24. У ряда животных и человека во время образования женских половых клеток (см.
Оогенез) М. останавливается на срок до нескольких лет и завершается только во время оплодотворения.
Первая фаза М. - профаза I, наиболее сложная и длительная (у человека 22,5, у лилии 8-10 суток), подразделяется на 5 стадий. Лептотена - стадия тонких нитей, когда хромосомы слабо спирализованы и наиболее длинны, видны утолщения - хромомеры. Зиготена - стадия начала попарного, бок о бок соединения (синапсиса, конъюгации) гомологичных хромосом; при этом гомологичные хромомеры взаимно притягиваются и выстраиваются строго друг против друга. Пахитена - стадия толстых нитей; гомологичные хромосомы стабильно соединены в пары - биваленты, число которых равно гаплоидному числу хромосом; под электронным микроскопом видна сложная ультраструктура в месте контакта двух гомологичных хромосом внутри бивалента: т. н. синаптонемальный комплекс, который начинает формироваться ещё в зиготене; в каждой хромосоме бивалента обнаруживаются 2 хроматиды; т. о., бивалент (тетрада, по старой терминологии) состоит из 4 гомологичных хроматид; на этой стадии происходит кроссинговер, осуществляющийся на молекулярном уровне; цитологические последствия его обнаруживаются на следующей стадии. Диплотена - стадия раздвоившихся нитей; гомологичные хромосомы начинают отталкиваться друг от друга, но оказываются связанными, обычно в 2-3 точках на бивалент, где видны хиазмы (перекресты хроматид) - цитологическое проявление кроссинговера. Диакинез - стадия отталкивания гомологичных хромосом, которые по-прежнему соединены в биваленты хиазмами, перемещающимися на концы хромосом (терминализация); хромосомы максимально коротки и толсты (за счёт спирализации) и образуют характерные фигуры: кресты, кольца и др. Следующая фаза М. - метафаза I, во время которой хиазмы ещё сохраняются; биваленты выстраиваются в средней части веретена деления клетки, ориентируясь
Центромерами гомологичных хромосом к противоположным полюсам веретена. В анафазе I гомологичные хромосомы с помощью нитей веретена расходятся к полюсам; при этом каждая хромосома пары может отойти к любому из двух полюсов, независимо от расхождения хромосом др. пар. Поэтому число возможных сочетаний при расхождении хромосом равно 2
n, где
n - число пар хромосом. В отличие от анафазы митоза, центромеры хромосом не расщепляются и продолжают скреплять 2 хроматиды в хромосоме, отходящей к полюсу. В телофазе I у каждого полюса начинается деспирализация хромосом и формирование дочерних ядер и клеток. Далее следует короткая интерфаза без редупликации ДНК - интеркинез, и начинается второе
деление М. Профаза II, метафаза II, анафаза II и телофаза II проходят быстро; при этом в конце метафазы II расщепляются центромеры, и в анафазе II расходятся к полюсам хроматиды каждой хромосомы. Эта классическая схема М. имеет исключения. Например, у растений рода ожика (Luzula) и насекомых семейства кокцид (Coccidae) в первом делении М. расходятся хроматиды, а во втором - гомологичные хромосомы, однако и в этих случаях в результате М. происходит редукция числа хромосом. Различия между сперматогенезом и оогенезом у животных и образованием микроспор и мегаспор у растений не отражаются на поведении хромосом в ходе М., хотя размеры и судьбы сестринских клеток оказываются разными. Известны аномалии М. У межвидовых гибридов все хромосомы, а у анеуплоидов (см.
Анеуплоидия) непарные хромосомы не способны конъюгировать и остаются в виде унивалентов; у автополиплоидов (см.
Автополиплоидия) образуются объединения более чем из 2 хромосом - т. н. мультиваленты. В каждом из этих случаев невозможна правильная редукция числа хромосом в анафазе I; образующиеся гаметы (с несбалансированными наборами хромосом) либо сами нежизнеспособны, либо дают нежизнеспособное или уродливое потомство (см.
Хромосомные болезни). Отсутствие хиазм (ахизматия) обычно приводит к тем же результатам, однако у самцов некоторых видов мух, в том числе у дрозофилы, хиазмы всегда отсутствуют, хотя гаметы образуются нормальные. Причины перехода клеток от деления путём митоза к М. в жизненном цикле каждого организма, а также молекулярные механизмы конъюгации гомологичных хромосом и кроссинговера исследуются.
Лит.: Соколов И. И., Цитологические основы полового размножения многоклеточных животных, в кн.: Руководство по цитологии, т. 2, М. - Л., 1966, с. 390-460; Райков И. Б., Карпология простейших, Л., 1967; Богданов Ю. Ф., Гомологичная конъюгация хромосом, в кн.: Успехи современной генетики, т. 3, М., 1971, с. 134-61; Swanson С. P., Cytology and cytogenetics, Englewood Cliffs, 1957; Rhoades М. М., Meiosis, в кн.: The cell. Biochemistry. Physiology. Morphology, v. 3, N. Y., 1961, p. 1-75; John B., Lewis K. R., The meiotic system, W. - N. Y., 1965; (Protoplasmatologia, Bd 6, fasc. 1); Westergaard М., Wettstein D. von, The Synaptinemal complex, в кн.: Annual Review of Genetics, v. 6, 1972, p. 71-110.
Ю. Ф. Богданов.
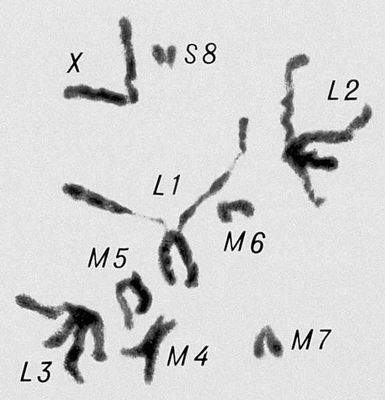
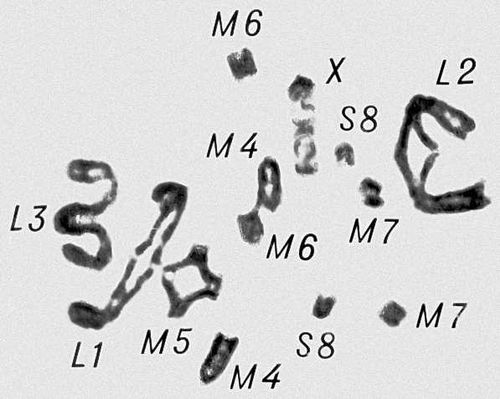
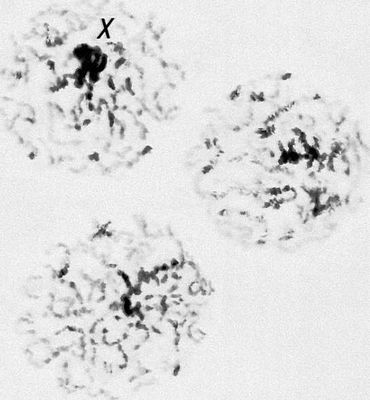
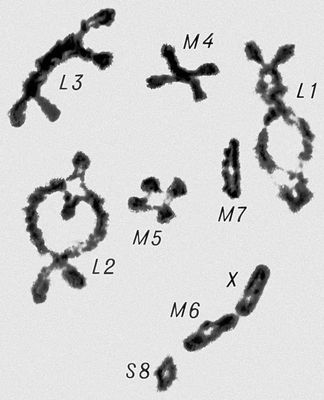
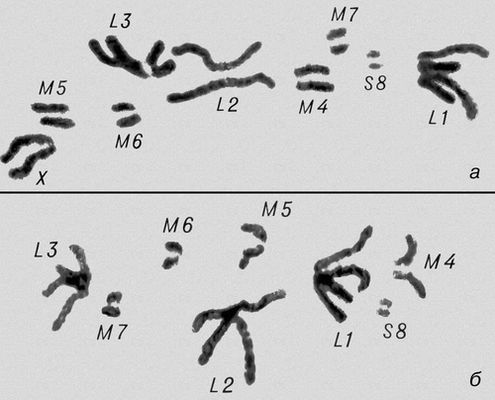
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Анафаза II; к полюсам расходятся хроматиды от каждой хромосомы.
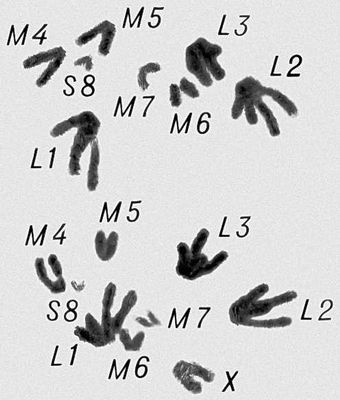
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Профаза II.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Метафаза I.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Интеркинез.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Диакинез; на этой стадии, так же как и в диплотене, легко сосчитать число бивалентов - их 8, и 1 унивалент; в каждом биваленте видны хиазмы.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Метафаза II; число хромосом гаплоидное, в каждой хромосоме видны 2 хроматиды (а - с Х-хромосомой, б - без неё).
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Анафаза I; гомологичные хромосомы расходятся к полюсам клетки, в каждой хромосоме видны 2 хроматиды.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Более поздняя диплотена, стрелками показаны хиазмы.
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Диплотена; начало отталкивания гомологичных хромосом друг от друга в каждом биваленте; гомологичные хромосомы остаются в контакте только в точках перекрёста - хиазмах (стрелка).
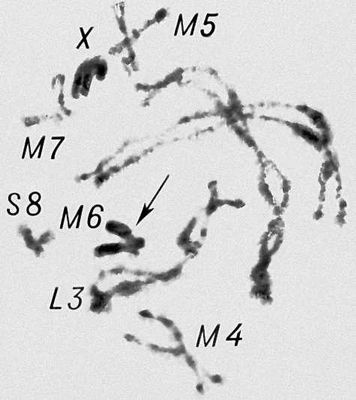
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Пахитена; все хромосомы конъюгируют попарно, образуя т. н. биваленты; Х-хромасома не имеет партнера для конъюгации и остается в виде унивалента (Х).
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Зиготена; двойные нити - сконъюгированные хромосомы (стрелка).
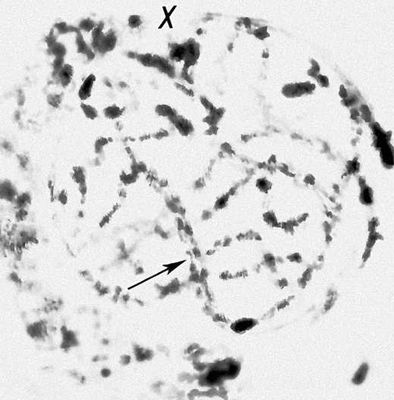
Морфология мейоза у самца кузнечика Chorthipus brunneus. Число хромосом - 17 (16 + Х): L - длинные хромосомы, М - средние, S - короткая, Х - Х-хромосома. Лептотена; тонкие одинарные нити - хромосомы (стрелка).
Схема мейоза: 1 - лептотена; 2 - зиготена; 3 - пахитена; 4 - диплотена; 5 - диакинез; 6 - метафаза I; 7 - анафаза I; 8 - телофаза I; 9 - интеркинез; 10 - метафаза II; 11 - анафаза II; 12 - телофаза II. Одна из двух гомологичных хромосом заштрихована, другая - белая. Обмен белыми и заштрихованными участками хромосом - результат кроссинговера. Маленькие белые кружки - центромеры, большой круг - контур ядра. В метафазе и анафазе обоих делений ядерная мембрана исчезает. В телофазе возникает снова. В метафазе и анафазе обоих делений стрелками показано направление растягивания и движения хромосом с помощью нитей веретена.